Оглавление
С.В.Наугольных
РАСТЕНИЯ ДАЛЕКОГО ПРОШЛОГО
Шаг Второй.
Морфогенез гомологичных органов
Эволюция спороношений хвощевидных; семейства
Tchernoviaceae и Equisetaceae.
(щелкните на рисунке, чтобы увеличить)
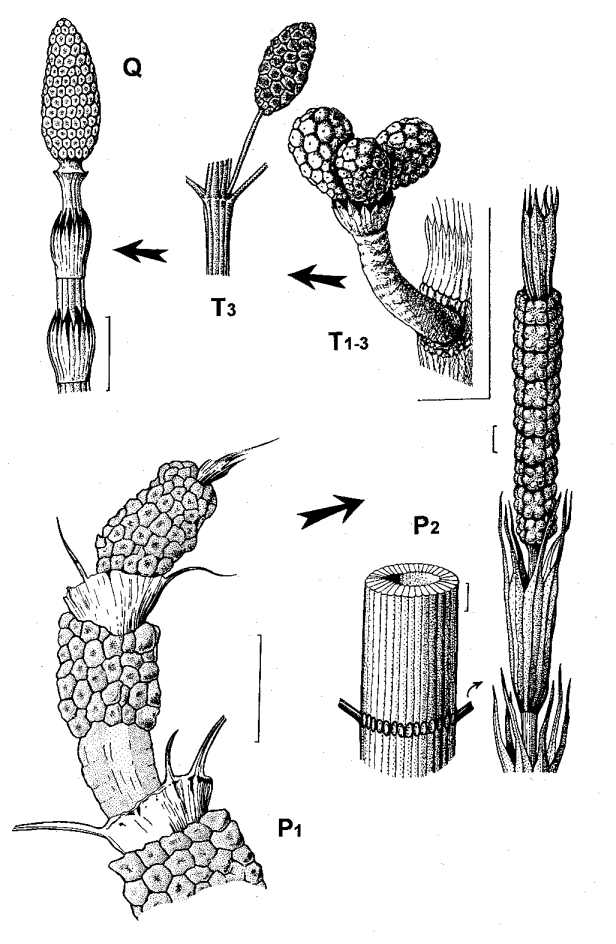
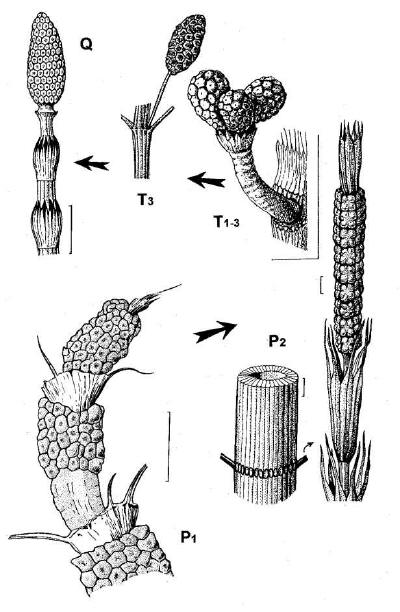
Рис. 2.
Морфогенетический тренд в эволюции представителей семейств Tchernoviaceae и
Equisetaceae; a – Equisetinostachys; б – Paracalamitina striata (слева –
фрагмент стебля с веточными рубцами, справа – терминальная фертильная зона,
“квазистробил”); в – Equisetites arenaceous (по Frentzen, 1934, с изм.); г –
Neocalamites aff. carrerei (по: Владимирович, 1958; с изм.); д – Equisetum
arvense. Длина масштабной линейки – 1 см (а, б, г-д), 2 мм (б).
|
Сидя на берегу
подтопленного заболоченного лесного озерца или речушки и глядя на густые
прибрежные заросли хвощей с длинными трескучими членистыми стеблями, легко
представить себя где-нибудь в конце палеозойской эры, в пермском периоде.
Близких и дальних родственников хвощей тогда было видимо-невидимо. Они, также
как и хвощи в современной растительности, в основном предпочитали селиться в
хорошо увлажненных местах и, судя по известным местонахождениям растительных
остатков этого возраста, нередко образовывали моновидовые сообщества.
Если открыть
практически любой из современных учебников ботаники на страницах, посвященных
геологической истории хвощей, то там мы прочитаем, что хвощи ведут свою
родословную от каламитов – гигантских споровых растений с членистыми стволами,
достигавшими 20 м в высоту, а то и более. Сейчас каламиты относятся к
самостоятельному семейству Calamitaceae или Calamostachyaceae, обособляемому в
особый порядок Calamitales или Calamostachyales (Мейен, 1987). С каламитами
современных хвощей сближает то, что продольные ребра, расположенные на побегах и
маркирующие положение проводящих тканей в побеге, и у тех, и у других чередуются
в узлах, в то время как у других палеозойских членистостебельных (например,
астерокаламитов или, как их еще называют, археокаламитов,
клинолистников-сфенофиллов, а также растений из семейства черновиевых, из
которых наиболее известен формальный род паракаламитес – Paracalamites Zalessky)
ребра проходят через узлы побега прямо, не чередуясь. Такое положение ребер на
побеге называют юкстапозицией. Из перечисленных выше групп от хвощей особенно
далеки сфенофилловые. Это очень своеобразная группа споровых растений,
заслуживающая специального разговора. От раннекаменноугольных астерокаламитовых,
судя по многим признакам, произошли черновиевые, также претендующие на роль
предков современных хвощей.
Спороношения
каламитов образуют компактные собрания (стробилы), внешне несколько напоминающие
стробилы современных хвощей. Однако если присмотреться к некоторым важным
деталям в строении хвощей и каламитов, выяснится, что между этими растениями
существует много различий. Главное из них заключается в том, что спорангиофоры
хвощей расположены на фертильной оси один за другим, примыкая друг к другу, а у
каламитов они находятся в пазухах видоизмененных чешуевидных листьев – брактей.
Спороношения другой группы позднепалеозойских членистостебельных, относившихся к
самостоятельному семейству черновиевых (Tchernoviaceae) в составе порядка
хвощевидных (Equisetales), имели существенно иное строение.
Черновиевые были
характерны, в основном, для карбона и перми Ангариды, большого материка,
располагавшегося в пределах современной северо-восточной Азии.
Из черновиевых в
качестве одного из наиболее типичных примеров можно рассмотреть род
Equisetinostachys.
Спороношения этого растения более сходны со стробилами современного хвоща, чем
спороношения каламитов. Щитки спорангиофоров на незрелых стробилах плотно
смыкаются своими краями. По мере созревания спорангиев, ножки щитков
спорангиофоров вырастают, удлиняются, а края щитков загибаются кверху, открывая
возможность спорам, высыпавшимся из созревших спорангиев, выйти наружу и
разнестись водой и ветром.
У
Equisetinostachys, промимо сходства в строении спорошений с современным хвощом,
наблюдаются и своеобразные черты строения. Спороношения Equisetinostachys
состояли не из единственного стробила, располагавшегося на верхушке побега, а из
нескольких фертильных зон, образовывавших сложную многоярусную конструкцию,
разделенную мутовками стерильных листьев. На верхушке фертильного побега
эквизетиностахиса располагался пучок листьев, указывающий на то, что верхушечные
меристемы этого побега (в отличие от стробилов современного хвоща) были
активными. Таким образом, спороношения Equisetinostachys не образовывали
настоящих специализированных стробилов, а располагались непосредственно на
побеге растения.
Какие же
палеозойские растения, каламиты или черновиевые, стояли в основании
филогенетической цепочки, ведущей к современным хвощам?
Перед тем, как
попытаться ответить на этот вопрос, надо подробнее поговорить о том, что нам об
этих растениях известно.
На реконструкциях
палеозойских ландшафтов древние членистостебельные нередко изображаются как
увеличенные во много раз копии современных хвощей. Надо сразу же оговориться,
что это неправильно. Конечно, как это уже отмечалось выше, и у современных
хвощей, и у их ископаемых родственников «габитуально» много общего. Прежде
всего, это моноподиально ветвящиеся побеги с ветвями и листьями, собранными в
мутовки. Но, по большому счету, на этом сходство во внешнем виде этих растений
заканчивается.
Формы роста
каламитов были реконструированы знаменитым немецким палеоботаником М.Хирмером
(Hirmer, 1927). Предложенные Хирмером реконструкции вошли во многие учебники
палеонтологии и ботаники. Исходя из особенностей расположения ветвей на побеге,
Хирмер выделил несколько подродов каламитов (иногда их используют в качестве
самостоятельных родов): Stylocalamites, Crucicalamites, Diplocalamites,
Calamitina. Позднее к реконструкциям каламитов, сделанных Хирмером, были
добавлены реконструкции еще нескольких представителей каламитов, основанные на
находках стволов с ветвями и даже спороношениями, а также
сравнительно-морфологических исследованиях (Noll, 2001).
Как выглядели при
жизни ангарские черновиевые долгое время оставалось загадкой. Было в общем-то
понятно, что, по меньшей мере, некоторые из этих растений были древовидными,
поскольку вместе с побегами, листьями и спороношениями черновиевых часто
встречались фрагменты крупных стволов диаметром до 10-15 сантиметров.
В представительном разрезе пермских отложений, расположенном по берегам (в
основном, на левобережье) реки Кожим в Печорском Приуралье мне удалось найти
крупный ствол членистостебельного, относившегося к черновиевым. К узлам ствола
прикреплялись побеги с листьями, а на концах некоторых из побегов сохранились
спороношения. Ствол уходил в твердую скальную породу, но после нескольких часов
работы молотком и зубилом удалось освободить ствол от породы на протяжении более
полуметра. Исходя из соотношения диаметра и скорости уменьшения толщины ствола
удалось вычислить приблизительную высоту растения, которая должна была превышать
3,5 метра. На основе этой находки была предложена первая графическая
реконструкция целого растения из семейства черновиевых (Naugolnykh, 2002). Ранее
были опубликованы реконструкции отдельных частей побегов (Рассказова, 1961) и
спороношений (Мейен, Меньшикова, 1983) некоторых других представителей
черновиевых.
У Paracalamitina striata спороношения, как и спороношения эквизетиностахиса,
сохраняли активность терминальных меристем: на верхушках фертильных зон, которые
можно условно назвать «квазистробилами», у них располагался все тот же хохолок
из стерильных листьев, но количество фертильных зон, по сравнению с
Equisetinostachys, сократилось до одной, в очень редких случаях – двух. Другое
пермское членистостебельное с намечающейся редукцией фертильных зон,
расположенных на боковых побегах, было описано С.В.Мейеном и Л.В.Меньшиковой
(1983) как Sendersonia matura Meyen et Mensh. из верхнепермских отложений
Кузбасса.
Один из эффективных
способов корректировки филогенетических гипотез, выдвигаемых палеонтологами,
заключается в изучении изменчивости и аберраций современных организмов, которые
иногда повторяют черты строения своих отдаленных предков. Происходит это потому,
что некоторые из «древних» генов сохраняются в кариотипе потомков в латентном,
«спящем» состоянии, а затем такие гены, нередко под воздействием неблагоприятных
факторов среды, могут активизироваться.
Летом 1998 г. в
приустьевой части Монастырского оврага (Ульяновская область, правый берег р.
Волга в 10 км выше по течению от г.Тетюши, у с. Монастырское) мне
посчастливилось найти аберрантные побеги современного хвоща зимующего -
Equisetum hyemale L., существенно отличающиеся от нормальных. В отличие от
обычных побегов, обнаруженные в Монастырском овраге экземпляры хвощей этого вида
несли стробилы, расположенные не на главном осевом побеге, а на нескольких
боковых побегах.
Длина собранных
побегов Equisetum hyemale составляла от 50 до 80 см при ширине стеблей 4 мм.
Длина междоузлий варьировала от 38 до 65 мм. На всем протяжении стебля (за
исключением самого верхнего узла) изученных экземпляров боковые ветви
отсутствовали. Отсутствие боковых ветвей - весьма характерный признак этого вида
хвощей. Самый верхний узел нес от двух до трех коротких боковых ветвей, каждая
из которых заканчивалась стробилом, что для этого вида (так же как и для
остальных современных хвощей) абсолютно нетипично. На основном побеге
располагался стробил, как правило, значительно лучше развитый, чем стробилы
боковых ветвей.
Такое расположение
стробилов на боковых ветвях характерно для мезозойских хвощей - представителей
родов Equisetites Sternberg, 1838 и Neocalamites Halle, 1908, причем известны
случаи, когда стробилы сидели на боковых ветвях по одному, как у описанного выше
E. hyemale (Neocalamites aff. carrerei (Zeiller) Halle из угленосных триасовых
отложений Челябинского бассейна; Владимирович, 1958; Основы палеонтологии,
1963). Сходное расположение стробилов (но не по одному, а по три стробила) на
укороченных боковых побегах отмечалось для Equisetites arenaceous (Jager) Schenk
(Frentzen, 1934; Kelber, van Konijnenburg-van Cittert, 1998).
Среди современных
Equisetum встречаются и другие аберрации. Известны экземпляры, стробилы которых
разделены на несколько фертильных зон, практически идентичных спороношениями
пермского рода Equisetinostachys, о котором уже шла речь выше. Иногда верхняя
фертильная зона может нести пучок стерильных листьев, аналогичных терминальному
"хохолку" у Equisetinostachys. Это явление называется пролиферацией.
У крупного
современного хвоща Equisetum telmateia Ehrh. описаны отклоняющиеся экземпляры, у
которых происходит развитие побега выше стробила, представляющего, по существу,
фертильную зону, расположенную на междоузлии (Kashyap, 1930; Tschudy, 1939).
Таких примеров, описанных в литературе, можно привести множество, хотя в
природе, разумеется, отклонения встречаются редко.
Форма стробиляции
на боковых побегах, сходная с Sendersonia matura Meyen et Mensh., описана у
Equisetum telmateia (Westwood, 1989), причем иногда на боковых побегах
образуются и фертильные зоны с пролиферацией.
Наличие у
современных хвощей аберраций, идентичных репродуктивным органам черновиевых
(Equisetinostachys, Sendersonia) указывает на прямую филогенетическую связь
семейства Equisetaceae с позднепалеозойскими ангарскими черновиевыми (семейство
Tchernoviaceae).
Таким образом,
можно констатировать следующий факт: у современного хвоща известны формы с
отклоняющейся от нормы морфологией, когда терминальная меристема стробила даже
после полного развития стробила остается активной и позволяет побегу расти
дальше (пролиферация). В этом случае такие аберрантные спороношения уже
практически ничем не отличаются от фертильных зон пермских Equisetinostachys или
Paracalamitina (в зависимости от количества повторно закладывающихся стробилов
на побеге). Кроме этого, известны аберрации, например, хвощ зимующий из
Монастырского оврага, при которых репродуктивные органы повторяют форму
триасовых хвощей.
Характерно, что
аберрантные спороношения с пролиферацией закладываются у современного Equisetum,
как правило, на боковых ветвях, а не на основном побеге, как нормальные
стробилы. По этому признаку такие отклоняющиеся хвощи также идентичны пермским
членистостебельным семейства Tchernoviaceae (роды Equisetinostachys,
Paracalamitina, Sendersonia). По существу дела, эта рекапитуляция примитивных
генов, находящихся в латентном состоянии, превращает современные хвощи с
аберрациями репродуктивных органов в уменьшенные во много раз копии их предков
из палеозойской эры.
Если выстроить
обсуждавшиеся выше формы в один ряд от древних видов к молодым, можно выявить
морфогенетический тренд, выражающийся в общей редукции и генеративных, и
вегетативных органов. Растения уменьшаются в размерах (от 3-4 метровых
паракаламитин до современных хвощей, большинство которых не превышают нескольких
десятков сантиметров в высоту). Репродуктивные органы сохраняются только на
верхушке побега, а многочисленные фертильные зоны редуцируются до единственной,
терминальная меристема которой теряет активность (при этом фертильная зона
превращается в стробил). По всей видимости, редукция сопровождалась ускорением
генеративных процессов: созревания репродуктивных органов и начала генеративной
активности (фертильности) растения в целом. Это давало предкам хвощей
преимущества в конкурентной борьбе и освоении подходящих экологических ниш.
Очень сходные
тенденции известны и во многих других группах относительно примитивных высших
растений: у папоротников, плауновидных, птеридоспермов, цикадовых и гинкговых.
Не исключено, что процессы редукции и олигомеризации имеют базовое значение в
эволюции споровых и голосеменных растений.
Эволюция фолиарных органов Peltaspermales
Закономерности морфогенеза листьев
пельтаспермовых птеридоспермов Ангариды
(щелкните на рисунке, чтобы увеличить)
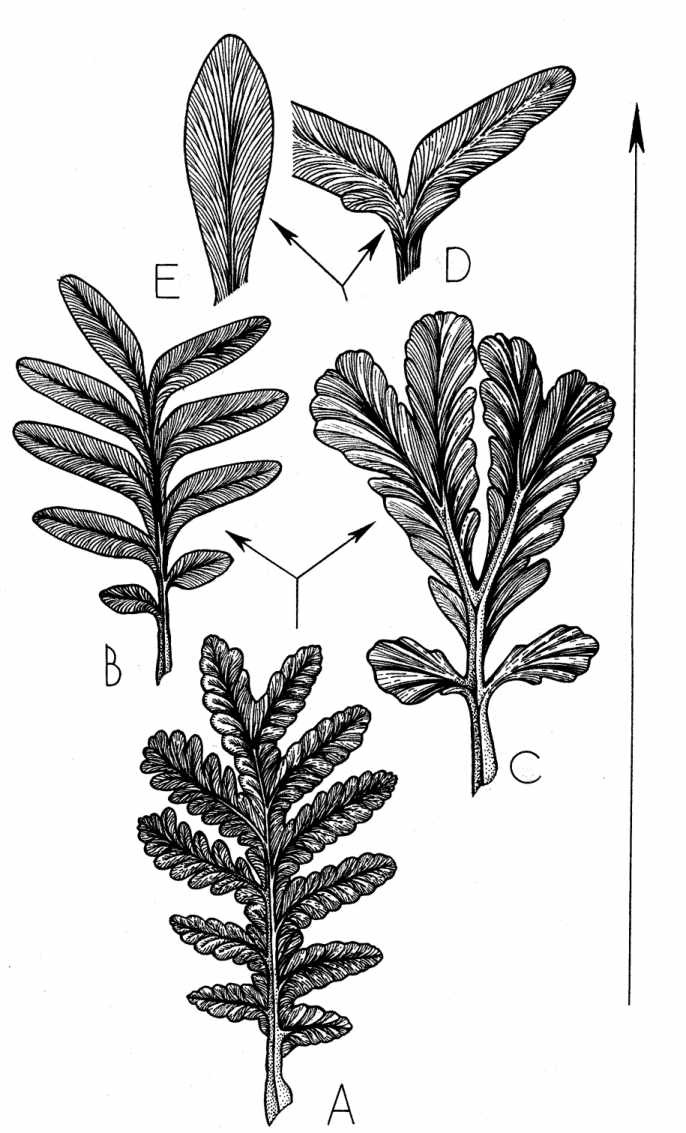
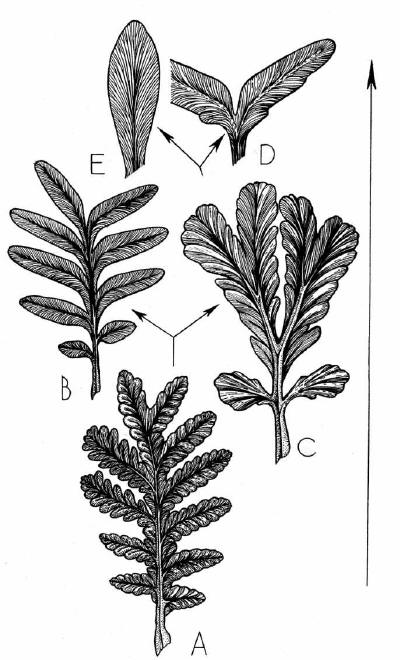
Рис. 3.
Морфогенетический тренд в развитии листьев пельтаспермовых: A- Rhachiphyllum
субангарского типа (артинский-кунгурский века), B - Psygmophyllum
(кунгурский-казанский века), C - Compsopteris (казанский век), D -
"Psygmophyllum" bifoliatum Zal. (татарский век), E - Pursongia (татарский
век). T - время. Подробное объяснение см. в тексте.
|
При работе с
остатками пермских пельтаспермовых птеридоспермов автор столкнулся с
необходимостью дать интерпретацию некоторым из макроморфологических
закономерностей, наблюдаемых при сравнительном анализе листьев этих растений.
Самым интересным из наблюдавшихся явлений было закономерное распределение
перышек (сегментов последнего порядка) сложноперистых листьев пельтаспермовых. В
подавляющем большинстве случаев, когда сохраняется целая или почти целая вайя,
верхние апикальные перья (сегменты предпоследнего порядка) всегда сильнее
надрезаны чем базальные. Апикальные перья несут перышки очень обособленные и
слабо срастающиеся основаниями. В проксимальной части той же вайи располагаются
перья с перышками, сильно сросшимися основаниями, боковыми краями, а иногда и
коалесцентно или когерентно слившиеся. Степень слитности перышек, таким образом,
закономерно увеличивается в проксимальном направлении вдоль оси (рахиса) вайи. В
самом основании вайи, около или вдоль черешковой части рахиса интеркалирующие
перышки сливаются иногда в сплошную полосу окаймления. Между наиболее широко
расставленными и обособленными перышками верхушки вайи и сильно сросшимися
перышками базальных перьев вайи наблюдается плавный переход.
Объяснить такую
высокую изменчивость перышек в пределах одного сложноперистого листа (вайи)
довольно трудно. Очевидно, что для одной вайи все факторы среды (увлажненность,
освещенность, эдафические условия) были одними и теми же. Вместе с тем,
привлечение к анализу возможности преобразования морфологических структур в
течение онтогенеза позволяет добиться логичной, внутренне непротиворечивой
интерпретации своеобразия архитектуры вайи пельтаспермовых птеридоспермов.
Физиологам растений
хорошо известно, что при различных условиях освещенности или увлажненности
растению выгодно иметь то сильно рассеченный, то более цельнокрайний лист. Вайи
со смешанным типом облиствения, видимо, были свойствены растениям, находившимся
в нейтральном физиологическом положении: у них в одинаковой степени были
проявлены оба признака. При смещении природных условий в ту или иную сторону
модус изменчивости листьев также менялся: при повышении влажности листья
становились более рассеченными, при повышении сухости перья
преобразовывались в когерентнолистные и уменьшались в размерах. Таким образом,
функционально нецелесообразное, на первый взгляд, сочетание разных типов перышек
на одной вайе может объясняться реальным проявлением потенциальных возможностей
растения регулировать эффективность транспирации за счет изменения степени
слитности перышек.
Модификацию
сложноперистых листьев птеридоспермов в зависимости от изменения факторов среды
легко пояснить в терминах олиго- и полимеризации. При более аридных условиях
модус изменчивости меняется в направлении олигомеризации, при более влажных - в
строну полимеризации.
Интересно, что
модификация
листьев птеридоспермов происходит закономерно и в масштабах
геологического времени (рис.). У представителей пельтаспермовых (порядок
Peltaspermales) наблюдается тренд в преобразовании сложноперистых
листьев
каллиптеридной морфологии (Rhachiphyllum) в более просто устроенные
простоперистые вайи Compsopteris или "четырехдельные" листья Psygmophyllum с
реликтами перистого рассечения. Филогенетически более молодые пельтаспермовые
(Tatarina, Pursongia, "Psygmophyllum" bifoliatum Zal.) имели простые
ланцетовидные или однократно бифуркирующие листья.
Очень симптоматично
совпадение тенденции к олигомеризации листьев пельтаспермовых в интервале от
конца ранней перми к началу триаса и общего иссушения климата в поздней перми,
причем пик аридизации приходился, очевидно, на самое начало триаса. Скорее
всего, упрощение морфологии листовой пластинки с одновременным уменьшением ее
размеров было "селекционным" ответом на аридизацию.
Читать дальше
Только для вас заказать цветы россия на выгодных условиях. Качественно. . По хорошей цене заказ цветов кино для всех желающих.